
Le piante sono organismi autotrofi che, letteralmente, non assimilano i nutrienti dall’esterno ma sintetizzano la propria biomassa carboniosa autonomamente. Per fare questo utilizzano in modo alquanto efficiente l’energia solare che irradia la crosta terrestre. Le piante si sono evolute convergendo verso il progressivo miglioramento dei processi fisici e chimici che sono alla base della fotosintesi in modo da rendere questa serie di reazioni quanto più efficiente ed utile possibile. Le piante, in virtù della fotosintesi, sono ecologicamente definite i “produttori primari” poiché sono gli organismi che, nella catena alimentare, per primi forniscono nutrienti organici, quali l’amido o la cellulosa, sintetizzati a partire da precursori non organici. Per la biosintesi di queste molecole è necessaria la presenza di una grande quantità di energia ed una organizzazione, a livello degli organi e delle singole cellule, che non può lasciare nulla al caso. Le piante soddisfano questi due requisiti perché risultano essere capaci di acquisire l’energia, sotto forma di fotone, e di processarla nelle strutture finemente organizzate che prendono il nome di cloroplasti.
La radiazione luminosa fornita dal sole può essere spiegata secondo due teorie: la teoria della luce come onda elettromagnetica e come corpuscolo. La luce ha, quindi, sia una proprietà di particella (fotone) sia una proprietà di onda, con le rispettive caratteristiche di frequenza e di lunghezza d’onda.
Se consideriamo la luce come corpuscolo dobbiamo comprendere che questo elemento può essere meglio definito come fotone. Il fotone possiede una determinata energia definita quanto che può essere espressa usando la legge di Planck:
![]()
Dove
h rappresenta la costante di Planck ed è pari a 6,626 · 10-34 J s
v rappresenta la frequenza
Vista la natura ondulatoria della luce vale anche il modello ondulatorio secondo il quale
![]()
Dove
c = Velocità della luce
v = Frequenza
λ = Lunghezza d’onda
Conoscendo le equazioni ondulatorie ed elettromagnetiche è possibile stabilire che
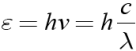
Osservando la precedente equazione è possibile stabilire che la radiazione a maggiore energia ha una minore lunghezza d’onda.
Il sole, come abbiamo già avuto modo di dire, rappresenta l’emettitore principale di energia luminosa. La componente di frequenza della radiazione solare non è omogenea e, in altre parole, la luce, intesa come serie di onde emesse dal sole, possiede differenti frequenze. Gli esseri umani percepiscono le frequenze dello spettro visibile e, ad ogni frequenza, si associa un determinato colore.

Illustrazione: Spettro visibile
Quando un fotone colpisce alcune strutture, quali quelle della clorofilla o degli altri pigmenti presenti nella pianta, può eccitare queste molecole portando un loro elettrone ad un livello energetico superiore. Questo fenomeno accade perché le molecole coinvolte nella fotosintesi, definite con il nome generico di pigmenti, possiedono una sensibilità a delle specifiche frequenze della luce.
Per rendere chiaro questo concetto supponiamo che in una cellula siano presenti una molecola di clorofilla “a”, il cui spettro di assorbimento è pari ad 680nm (luce rossa), ed una molecola di clorofilla “b” che assorbe a 480nm (luce verde). Se forniamo a questa ipotetica cellula soltanto della luce rossa sarà la clorofilla “a” ad avere il maggior beneficio poiché la frequenza della luce coincide con il valore ottimale di assorbimento per il pigmento in esame. Se, invece, forniamo luce verde sarà la clorofilla di tipo “b” ad essere favorita poiché la radiazione luminosa fornita coincide con il valore ottimale di assorbimento.
Questo spiega perché nella fotosintesi non è una sola clorofilla ad essere coinvolta nel processo ma diversi tipi di clorofilla uniti a numerosi altri tipi di pigmenti definiti, globalmente, con il termine di pigmenti accessori. In questo modo la luce solare, che presenta differenti frequenze corrispondenti ai diversi colori visibili, può essere captata non in una sua piccola fetta, ad esempio nella cromaticità del rosso, ma nella sua interezza.
Non tutta la pianta opera la fotosintesi clorofilliana. Un prerequisito chiave per lo svolgimento di questo processo è quello per il quale le cellule fotosintetiche devono essere esposte ad un sufficiente quantitativo di luce. Per questo motivo le radici, che affondano nella terra, non possono essere eleggibili per compiere le reazioni fotosintetiche a differenza della foglia e del fusto che, di norma, sono parti esposte alla radiazione solare per un tempo sufficientemente ampio.
L’organulo subcellulare nel quale si svolge la fotosintesi è il cloroplasto. Questo presenta una doppia membrana biologica ed una organizzazione ultrastrutturale caratteristica osservabile al microscopio elettronico. Nello stroma, che è lo spazio interno alla membrana del cloroplasto, sono presenti i tilacoidi ovvero compartimentazioni interne che non comunicano direttamente con l’esterno. I tilacoidi sono organizzati in modo da rimanere impilati, ovvero uno sopra all’altro, formando le cosiddette grana dei tilacoidi. Le varie grana dei tilacoidi sono unite dalle lamelle stromatiche.
I pigmenti
| Nome | Presenza | Assorbimento |
|---|---|---|
| Clorofilla “a” | PS-II (P680)
PS-II (antenna prossimale)PS-II (antenna distale)PS-I (P700) |
680nm(rosso)700nm (rosso) nel PS-I |
| Clorofilla “b” | PS-II (antenna distale) | 480nm(verde) |
| Carotenoidi | PS-I | 500nm(blu) |
Tabella: Picco di assorbimento e localizzazione dei principali pigmenti fotosintetici
Abbiamo già osservato che i fotoni colpiscono gli organi fotosintetici in strutture ben determinate che prendono il nome di pigmenti. Dal punto di vista biochimico i pigmenti sono molecole molto eterogenee tra loro ma c’è un elemento in comune che è dato dalla presenza di atomi, o di legami, particolari che riescono a captare il fotone, eccitandosi, ad una determinata lunghezza d’onda. Per quanto riguarda l’assorbimento è bene chiarire che ogni pigmento possiede un proprio optimum di frequenza che corrisponde ad un determinato colore dello spettro visibile nel quale l’efficienza di assorbimento, ovvero la capacità di tramutare l’energia quantica in energia capace di eccitare il pigmento, risulta ottimale. La clorofilla “a” assorbe ad una lunghezza d’onda di 680nm che corrisponde al colore rosso, la clorofilla “b” assorbe a 480nm (verde) mentre i carotenoidi hanno un picco di assorbimento pari a 500nm (blu).
La fase luminosa della fotosintesi
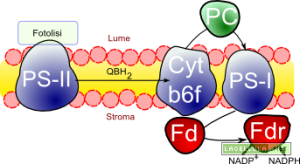
Illustrazione: Schematizzazione della fotosintesi clorofilliana
I pigmenti, dunque, sono disposti in modo tale da ottenere quanta più energia possibile per singola unità di superficie. L’organizzazione di questi pigmenti prende il nome di complesso antenna e rafforza l’idea che queste strutture servano da ricevitori eterogenei dell’energia presentata sotto forma di quanto. Il complesso antenna convoglia, mediante il fenomeno della risonanza, l’energia verso un centro di reazione che sarà il mezzo grazie al quale un elettrone, proveniente dall’ossidazione dell’acqua, verrà ceduto alla catena di trasporto.
Il fotosistema non è altro che l’unione del complesso antenna, con tutti i suoi pigmenti, ed il centro di reazione che possiede una particolare clorofilla di tipo “a”. È questa clorofilla che permette la trasduzione dell’energia da fisica a chimica in una serie di reazione di ossidoriduzione che hanno il compito di trasportare l’elettrone, generando quindi un flusso elettronico, verso il destino finale della fotosintesi: la produzione di equivalenti di riduzione sotto forma di NADPH e di “energia chimica” in forma di ATP o in altre reazioni enzimatiche di ossidazione. Per rendere possibile la sintesi di queste due molecole, nel cloroplasto, lavorano due fotosistemi definiti fotosistema I e fotosistema II abbreviati con i loro rispettivi acronimi PS-I e PS-II.
Fotosistema II
Il complesso antenna del fotosistema II capta i fotoni grazie al lavoro svolto dai pigmenti in esso presenti. I vari pigmenti convogliano verso il centro di reazione P680 il cui numero nella sigla indica che la clorofilla del centro di reazione assorbe la radiazione luminosa a 680nm corrispondenti al colore rosso. È importante comprendere che la luce, nella sua componente cromatica rossa, può colpire direttamente il centro di reazione oppure colpire un pigmento ad esso associato del complesso antenna ed essere trasferita per risonanza al P680.

Illustrazione: Flusso di elettroni del fotosistema II.
Il P680 è formato da un dimero intrecciato al cui interno figura esclusivamente clorofilla “a”. Dal punto di vista strutturale il centro di reazione del fotosistema II è formato da due polipeptidi denominati D1 e D2. In D1 si trovano il residuo Z, la feofitina ed il chinone QB. In D2 troviamo il chinone QA.
Abbiamo illustrato i pigmenti delle antenne dei fotosistemi, adesso ci occuperemo di analizzarli attentamente. Attorno al P680 sono presenti numerosi pigmenti di clorofilla “a” organizzati nel complesso antenna prossimale. I pigmenti di questo complesso hanno il compito di ampliare il raggio di cattura dei fotoni sulla lunghezza dei 680nm nel senso che possono, grazie alla loro presenza, catturare per unità di superficie molti più fotoni rispetto a quelli catturabili dal solo centro di reazione. Il complesso antenna prossimale assieme al centro di reazione, forma il cosiddetto “core”.
Il sistema di antenne prossimali si può suddividere in LHC-2 ed ACP-2. Il primo è un elemento fisso e contiene sia clorofille “a” che clorofille “b” mentre il secondo è un elemento mobile e contiene clorofille “a”, clorofille “b”. La differenza tra LHC-2 ed ACP-2 può essere determinata dal rapporto, in termini di quantità, tra clorofille di tipo “a” e clorofille di tipo “b”.
| Zona | Tipo |
|---|---|
| Antenne prossimali | Esclusivamente clorofille di tipo “a” |
| LHC II (distale) | Clorofille di tipo “a” e “b” nel rapporto di 2:1 |
| ACP II (distale) | Clorofille di tipo “a” e “b” nel rapporto di 1:2 oltre a varie xantofille |
Tabella: Pigmenti e clorofille nel fotosistema II e loro rapporti. Le clorofille b sono all’esterno del sistema (antenna distale) perché assorbono a lunghezze d’onda inferiori (luce blu). Siccome per ogni passaggio la lunghezza aumenta in questo modo si arriva agevolmente alla lunghezza d’onda di 680nm.
Nel fotosistema II si assiste ad un particolare fenomeno biologico: l’ossidazione dell’acqua. Lo stato di ossidazione dell’ossigeno nella molecola dell’acqua è pari a -2 mentre l’ossigeno molecolare ha numero di ossidazione pari a 0, per cui l’ossigeno, durante il processo, subisce ossidazione. I meccanismi con i quali l’acqua viene ossidata non sono ancora del tutto chiari ma è ipotizzabile la presenza di un sistema ciclico che sfrutta l’energia fornita dai fotoni per ossidare l’acqua. È importante comprendere che la vita aerobica è resa possibile grazie alla caratteristica dell’apparato fotosintetico vegetale di formare ossigeno partendo dall’acqua.
L’equazione generale della fotolisi dell’acqua è la seguente
![]()
Servono due molecole di acqua per formare una molecola di ossigeno, quattro protoni e quattro elettroni. I protoni permangono nella faccia interna del tilacoide definita lume e, come vedremo, contribuiscono alla acidificazione di questa zona. Gli elettroni, invece, vengono captati da particolari molecole e condotti lungo una catena di trasporto.
Trasporto degli elettroni dal centro di reazione
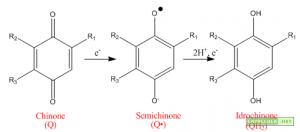
Illustrazione : Riduzione del chinone
L’idrochinone (QBH2) si dissocia dal complesso del fotosistema II e lungo la membrana tilacoidale incontra il citocromo b6f che è un complesso proteico che presenta numerosi gruppi prosteici. L’idrochinone B (QBH2) ha una fisiologica necessità di ossidarsi a chinone, tornando dunque al proprio stadio iniziale. Questo poiché la cellula vegetale non può permettersi di sintetizzare un elevato quantitativo di questo trasportatore, ed è proprio a livello del citocromo b6f che il QBH2 si ripristina ossidandosi
Citocromo b6f
Il citocromo b6f rappresenta la connessione tra il fotosistema II ed il fotosistema I; è da questo complesso che transitano gli elettroni che serviranno per ridurre il NADP a NADPH oppure verranno utilizzati per alcune reazioni enzimatiche.
Dal punto di vista strutturale il citocromo b6f è costituito da due dimeri, ognuno dei quali è formato da 4 grandi subunità e 4 piccole subunità. La parte estrinseca del citocromo è localizzata nel lume. Internamente ogni monomero possiede diverse unità di eme e un cluster che contiene due atomi di ferro e due di zolfo, letterariamente identificato come proteina ferro zolfo.
In un primo momento l’idrochinone QBH2 trasferisce la propria coppia di elettroni al complesso del citocromo: un elettrone viene donato alla proteina ferro zolfo ed un altro elettrone al citocromo b. La proteina ferro zolfo, a sua volta, cede l’elettrone al citocromo f che lo trasferisce alla plastocianina. L’altro elettrone viene ceduto al citocromo b che lo veicola su un chinone B formando una specie radicalica QB•.
A questo punto esiste una molecola radicalica che, in quanto tale, è potenzialmente dannosa per la cellula vegetale. Per rimediare a questa situazione, quando una ulteriore molecola di idrochinone arriva al citocromo, un nuovo elettrone viene trasferito alla proteina ferro-zolfo mentre un altro elettrone termina la riduzione della specie radicalica che si era appena formata mettendo a disposizione una nuova molecola di idrochinone. In altre parole un chinone QB dapprima viene ridotto a radicale QB• e successivamente viene ridotto a QBH2.
Fotosistema I
Nel fotosistema II gli elettroni provenienti dall’ossidazione dell’acqua vengono convogliati lungo una catena di trasporto che passa dal citocromo b6f e, attraverso la plastocianina, arriva al centro di reazione del fotosistema I.
Il fotosistema I (o PS-I) è sempre espresso nelle cellule vegetali a differenza del fotosistema II che può non essere presente. Il PS-I è formato da un centro di reazione (P700) circondato da un complesso antenna e da un una serie di trasportatori che hanno il compito di trasferire gli elettroni fino alla ferrodossina.
| Zona | Tipo |
|---|---|
| Antenna del fotosistema I | Contiene clorofille “a” e “b”, xantofille, beta-carotene |
Tabella: Pigmenti del fotosistema I
Al pari di quanto accade per il fotosistema II anche il fotosistema I possiede un complesso di pigmenti antenna che, assorbendo la radiazione luminosa, rendono possibile l’eccitazione di una particolare clorofilla in esso presente. La clorofilla trasferisce gli elettroni ad una serie di altre molecole tra cui figurano un fillochinone ed una serie di molecole ferro-zolfo. La penultima molecola è la ferrodossina che cede gli elettroni ad una flavoproteina chiamata ferrodossina-NADP-reduttasi che rende possibile la riduzione del NADP a NADPH.
Ferrodossina e trasporto alternativo di elettroni
| Accettore | Ruolo |
|---|---|
| Ferrodossina-NADPH-reduttasi | Riduzione del NADP a NADPH |
| Nitrogenasi | Riduzione dell’azoto atmosferico (N2) ad ammonio (NH4+) |
| Chinone B | Nella fotosintesi ciclica l’elettrone viene trasferito al QB per evitare fotodanno. |
| Nitrito reduttasi | Riduzione mitocondriale del nitrito in ammonio. |
Tabella: Trasporti alternativi degli elettroni dal PS-I
Gli elettroni che arrivano alla ferrodossina possono seguire delle vie alternative rispetto a quelle enzimatiche che portano alla riduzione del NADP. Il PS-I, difatti, “alimenta” numerose reazioni enzimatiche che possono essere compiute proprio in virtù dell’energia che transita dalla ferrodossina.
Fotosintesi Ciclica
Il fotosistema I può operare indipendentemente dal fotosistema II e questa peculiarità viene a manifestarsi nelle cellule che hanno più bisogno di energia sotto forma di ATP quali, ad esempio, le cellule di guardia degli stomi o in quelle cellule dove la concentrazione di ossigeno può risultare dannosa per alcune vie metaboliche, ad esempio nelle eterocisti presenti in alcuni batteri fissatori di azoto che presentano cloroplasti mancanti del PS-II.
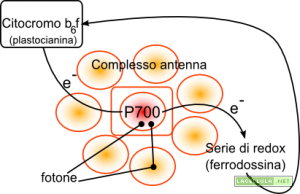
Illustrazione: Schema della fotosintesi ciclica
La ferrodossina può cedere l’elettrone anziché all’enzima ferrodossina-NADP-reduttasi nuovamente al citocromo b6f e, in questo modo, viene messa in opera la cosiddetta fotosintesi ciclica dove il fotosistema I è di fatto indipendente dal fotosistema II che può non essere affatto presente. Nella fotosintesi ciclica non si produce ossigeno, che viene prodotto grazie al complesso di ossidazione dell’acqua che troviamo nel fotosistema II, ma soltanto ATP perché il citocromo b6f è responsabile della traslocazione di protoni dallo stroma al lume tilacoidale. Nella fotosintesi ciclica, inoltre, proprio perché la ferrodossina non trasferisce i propri elettroni all’enzima ferrodossina-NADP-reduttasi non è possibile la sintesi di equivalenti di riduzione sotto forma di NADPH.
Produzione di ATP nella fotosintesi ed il bilancio energetico
A questo punto è possibile fare un bilancio stechiometrico della fase luminosa della fotosintesi. Per portare una molecola di NADP alla sua forma ridotta (NADPH) sono necessari dieci fotoni. Quattro fotoni servono per la fotolisi dell’acqua mentre, altri quattro, servono per promuovere due elettroni dal PS-II verso la catena di trasporto ed ulteriori due fotoni sono necessari per ripristinare il centro di reazione P700 del PS-I.