
L’acqua viene assorbita dalle radici per essere trasportata, mediante una serie di processi, verso ogni periferia della pianta. Sappiamo che l’acqua è una molecola essenziale per lo sviluppo dell’organismo vegetale ma, volendo essere più specifici, qual’è il destino dell’acqua una volta che questa viene assorbita dalle radici?
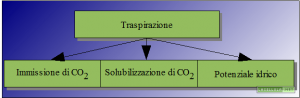
Disegno 1: Effetti della traspirazione
I motivi per i quali la pianta necessita di acqua, e di conseguenza, di un terreno ben idratato sono diversi ed abbracciano fenomeni chimici e fisici. Nella fotosintesi clorofilliana l’acqua e l’anidride carbonica sono le molecole inorganiche che servono per la formazione del glucosio. L’acqua può anche essere considerata come una molecola “di struttura” in quanto garantisce la turgidità delle cellule non lignificate e, nel complesso, rappresenta una grande aliquota del peso totale della pianta. L’acqua, inoltre, è presente nel citoplasma e serve per le reazione biochimiche ad esempio di idratazione sia perché gli enzimi lavorano, per la maggior parte, in soluzione acquosa. Nonostante tutto la pianta ha un “canale aperto” dal quale una discreta quantità di questo prezioso fluido viene persa sotto forma di vapore acqueo nel fenomeno della traspirazione.
A livello della foglia, specialmente nella faccia inferiore, sono presenti dei piccoli canali che mettono in comunicazione il mesofillo con l’ambiente esterno. Le aperture appena citate vengono definite stomi e attraverso questi minuscoli pori l’acqua fuoriesce dalla pianta e si disperde verso l’esterno sotto forma di vapore acqueo.
Se l’acqua è così importante, ed effettivamente lo è, per quale motivo la pianta la lascia andar via così facilmente? Per rispondere a questa domanda dobbiamo tener conto che l’organismo vegetale ha bisogno di assumere l’anidride carbonica dall’esterno.
Funzione della traspirazione
Abbiamo appena ricordato che, nella fotosintesi, si sintetizza lo zucchero glucosio a partire da due molecole inorganiche tra cui l’anidride carbonica. Questa molecola viene presa dall’atmosfera sotto forma di gas e, per questo motivo, la pianta ha la necessità di lasciare gli stomi aperti per acquisire CO2. L’acqua, in questo modo, viene persa ma contemporaneamente la pianta può acquistare anidride carbonica da usare nei processi fotosintetici.
La fotosintesi avviene nel cloroplasto che è un organulo cellulare delimitato da una doppia membrana biologica. L’anidride carbonica non riesce ad oltrepassare le membrane del cloroplasto direttamente in forma gassosa ma necessita di essere disciolta in acqua. A livello dello stoma la presenza del vapore acqueo traspirato rende possibile l’ingresso dell’anidride carbonica in forma disciolta.
Un’altra spiegazione che giustifica la traspirazione è correlata al meccanismo di trasporto dell’acqua dalla radice al meristema ed alle foglie. Abbiamo detto che l’acqua si sposta in base al potenziale di pressione tra due compartimenti. Se noi operiamo una estrema semplificazione della pianta e la consideriamo come l’insieme di due compartimenti unici, ovvero radice e foglia, proprio perché il potenziale idrico diventa sempre più negativo man mano che aumenta la concentrazione del soluto, ovvero man mano che diminuisce il quantitativo di acqua, possiamo capire che se l’acqua rimanesse in quantità notevole nel compartimento fogliare aumenterebbe il potenziale idrico. In altre parole il potenziale idrico tra radice e foglia, in un tempo più o meno lungo, arriverebbe all’equilibrio e l’acqua non potrebbe più risalire causando la sofferenza dei tessuti vegetali.
Regolazione della traspirazione dagli stomi
Per impedire l’eccessiva traspirazione la foglia è ricoperta dalla cuticola, che è uno strato ceroso altamente impermeabile che garantisce una ridottissima perdita di vapore acqueo. Lo strato ceroso è interrotto dagli stomi che sono dei piccoli pori più o meno uniformemente distribuiti lungo la superficie fogliare. Nei momenti di necessità, ovvero quando la pianta ha bisogno di una maggiore o minore presenza di anidride carbonica, vengono aperti o chiusi gli stomi che, in un certo senso, servono da ponte tra atmosfera e tessuto vegetale. La cuticola non può controllare il flusso di vapore acqueo/anidride carbonica perché la sua funzione primaria è quella di limitare l’evaporazione dell’acqua per cui la presenza degli stomi, che interrompono la continuità della cuticola, si rende necessaria per garantire un apporto corretto di gas alla cellula.
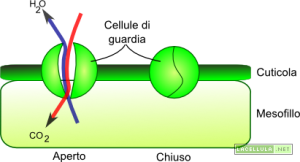
Illustrazione 13: Stomi e cellule di guardia
Lo stoma non è statico ma può regolare la propria forma in modo tale da consentire, o impedire, la fuoriuscita d’acqua. Le cellule di guardia che lo formano possono riempirsi di acqua aumentando il proprio turgore, e aprire lo stoma, o viceversa perdere acqua per chiudere lo stoma. La regolazione di aperture e chiusura degli stomi è finemente regolata da meccanismi biochimici e, vedremo, è garantita dalla particolare morfologia delle cellule di guardia che formano queste aperture.
Meccanismo di apertura degli stomi
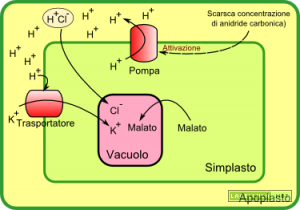
Illustrazione: Meccanismo di apertura degli stomi
Quando i sensori della CO2 captano una bassa concentrazione della molecola attivano le pompe protoniche che estrudono dei protoni portandoli dal citoplasma all’apoplasto.
Tra apoplasto e simplasto, per via dell’eccesso di cariche positive nel primo e negative nel secondo, è ipotizzabile che si instauri una differenza di potenziale che agisce su specifici trasportatori dello ione potassio che, una volta trasportato dentro il simplasto, si accumula nel vacuolo. Altri trasportatori, definiti trasportatori simporto, associano il rientro del protone ad una specie carica negativamente (come il cloro) che, altrimenti, proprio per via della carica, non avrebbe potuto attraversare la membrana. In questo modo specie a carattere ionico possono arrivare fino al vacuolo e, secondo l’equazione che calcola il potenziale idrico, contribuiscono ad abbassare il potenziale dell’acqua.
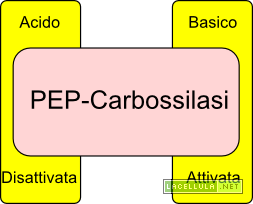
Abbiamo visto che l’attività di alcuni ioni contribuisce a far entrare acqua in quanto si assiste ad una diminuzione del potenziale idrico. Se torniamo un attimo all’equazione del potenziale idrico notiamo che questo è calcolato anche in base al potenziale di matrice che è la misura dell’attività di alcune molecole organiche. Il malato, ovvero l’anione dell’acido malico, è una molecola a quattro atomi di carbonio la cui sintesi citosolica avviene ad opera dell’enzima PEP-carbossilasi particolarmente attivo in fase di apertura degli stomi; nello specifico l’alcalinizzazione del citoplasma comporta l’attivazione dell’enzima.
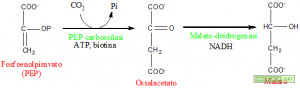
Illustrazione: Biosintesi del malato.
La PEP-carbossilasi addiziona al fosfoenolpiruvato (PEP) una molecola di anidride carbonica per formare l’ossalacetato. La malato deidrogenasi, un enzima NADH dipendente, riduce l’ossalacetato in malato che entra, mediante un trasportatore, nel vacuolo contribuendo all’abbassamento del potenziale idrico.
Meccanismo di chiusura degli stomi
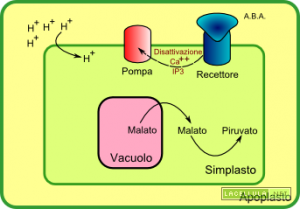
Illustrazione: Meccanismo di chiusura degli stomi
L’organismo vegetale si è evoluto nella direzione della segnalazione ormonale e, per quanto riguarda la perdita d’acqua, la molecola di acido abscissico (ABA) rappresenta un potente modulatore della chiusura degli stomi.
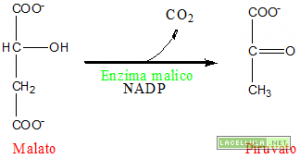
Illustrazione: Sintesi del piruvato ad opera dell’enzima malico
Quando l’acido abscissico colpisce un recettore posto nella parete cellulare, coadiuvato dall’azione degli ioni calcio (Ca++), disattiva la pompa protonica. Il gradiente esterno di protoni viene meno in quanto queste particelle entrano nuovamente nel simplasto. La perdita di alcalinità del simplasto disattiva la PEP-Carbossilasi che, di conseguenza, non produrrà più ossalacetato ed in risposta il malato vacuolare torna nel citoplasma. L’acidificazione del citoplasma, inoltre, disattiva la PEP-carbossilasi e porta all’attivazione dell’enzima malico che processa il malato ossidandolo a piruvato e trasferendo equivalenti di riduzione al NAD, riducendolo. Inoltre si inverte il canale di trasporto dell’anione cloro (Cl–).
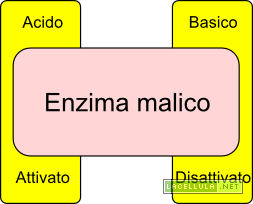
Illustrazione: Regolazione dell’enzima malico in base al pH citoplasmatico
Abbiamo detto che gli ioni calcio disattivano la pompa protonica: ma da dove provengono? Gli ioni calcio vengono rilasciati dal reticolo endoplasmatico in seguito all’azione dell’ormone ABA che, a seguito di alcune attivazioni e reazioni, media il rilascio degli ioni.
È da notare che, dal punto di vista del potenziale idrico, nell’apertura degli stomi e nella chiusura degli stomi si verificano due fenomeni totalmente opposti ma finemente regolati.
C’è un caso nel quale gli stomi non si chiudono in base alla concentrazione di CO2 o di ABA: la chiusura idropassiva. La chiusura idropassiva degli stomi avviene quando la pianta è in una fase di stress idrico talmente pesante da far diminuire l’aliquota di acqua presente a livello delle cellule di guardia. La chiusura idropassiva non è controllabile né dall’ABA né da nessun altro meccanismo e, a differenza del controllo ormonale che è indotto, è un fenomeno del tutto fisico.